Se han establecido 36 parcelas permanentes de 1 hectárea en la región Amazónica (Figura 5)
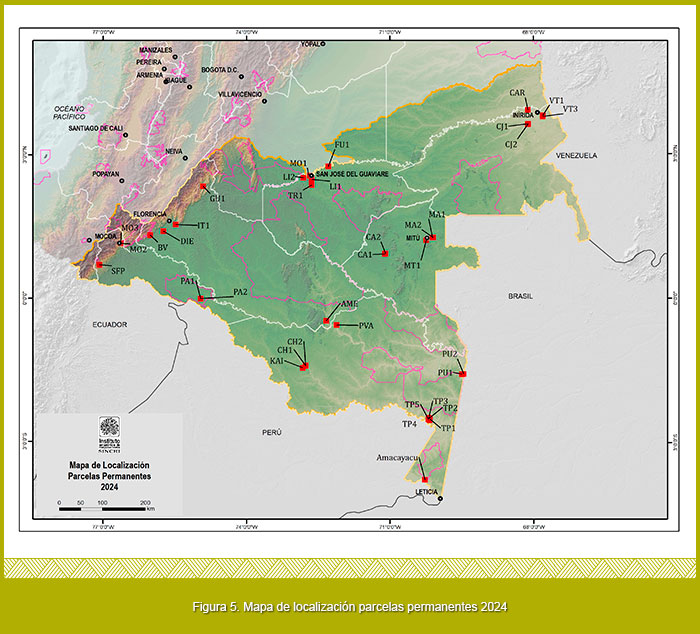
En total se tienen monitoreados 21.830 árboles correspondientes a 2.126 especies, 457 géneros y 107 familias.
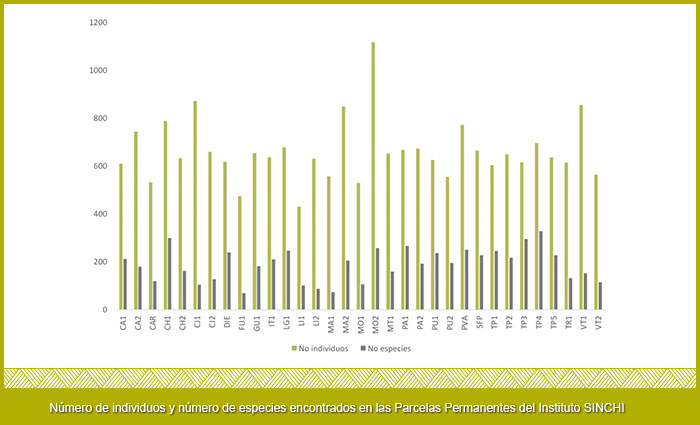
Las parcelas con mayor número de individuos son MO2 con 1116 individuos, seguida de CJ1 con (871 ind.), por otro lado las parcelas con menor número de individuos fueron LI1 (430 ind.) y FU1 (474 ind.); a su vez las parcelas más diversas fueron Tarapacá 4 con 328 especies seguida de CH1 con 299 especies, las parcelas menos diversas fueron FU1 con 69 especies y MA1 con 73 especies (Figura 6).
Resultados Parcelas 1 ha
2024
Castaño et al 2024 https://doi.org/10.1111/ecog.06693
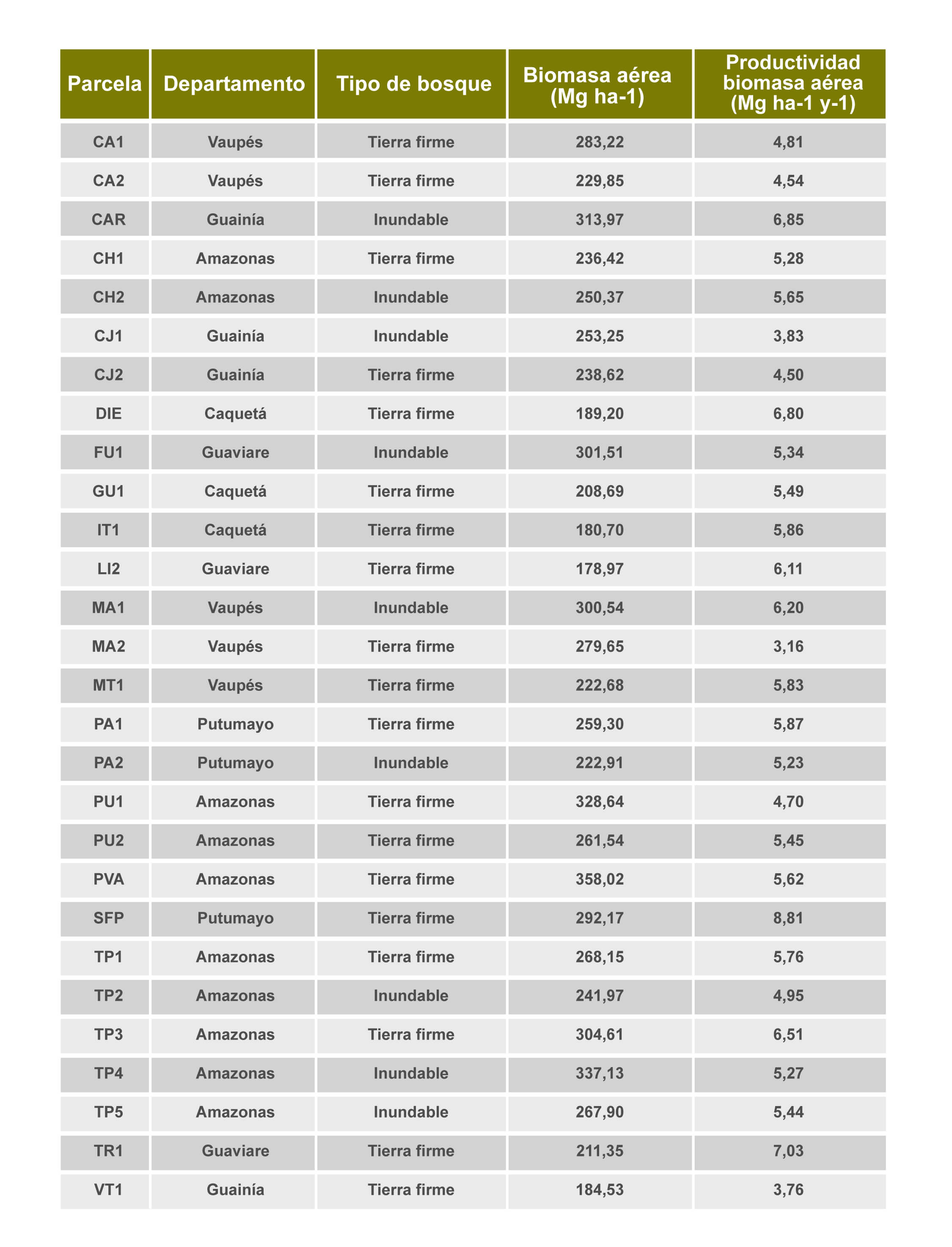
Evaluamos la importancia relativa de los factores abióticos y bióticos en la configuración de las existencias y los flujos de biomasa forestal (biomasa aérea) (https://doi.org/10.1111/ecog.06693). Utilizamos el conjunto de datos de la Red de Parcelas del Instituto SINCHI y parcelas de colegas de la Universidad Nacional de Colombia, sede Medellín y la Universidad de los Andes, en donde recopilamos información de la dinámica de 39 parcelas de 1 hectárea (ha) ubicadas en bosques maduros inundables y de tierra firme de la Amazonía y Orinoquía en Colombia.
Evaluamos la importancia del clima, la fertilidad del suelo y las inundaciones, así como la diversidad taxonómica y filogenética de los árboles y las propiedades estructurales de los bosques, frente a la determinación de las reservas de biomasa aérea (AGB por sus siglas en inglés; Mg ha−1) y la productividad de biomasa aérea (AWP por sus siglas en inglés; Mg ha−1 año−1). Utilizamos un método de inferencia multimodelo proveniente de la teoría de la información para identificar los principales motores frente a la biomasa y su productividad. Encontramos que características estructurales del bosque, como el número de árboles con un diámetro a la altura del pecho ≥ 70 cm, y la densidad de la madera, son los principales impulsores de la variación en las reservas de biomasa aérea. Sin embargo, la diversidad taxonómica también contribuye a la biomasa aérea, los árboles más grandes son de ciertas familias de plantas (Castaño et al 2024 https://doi.org/10.1111/ecog.06693)
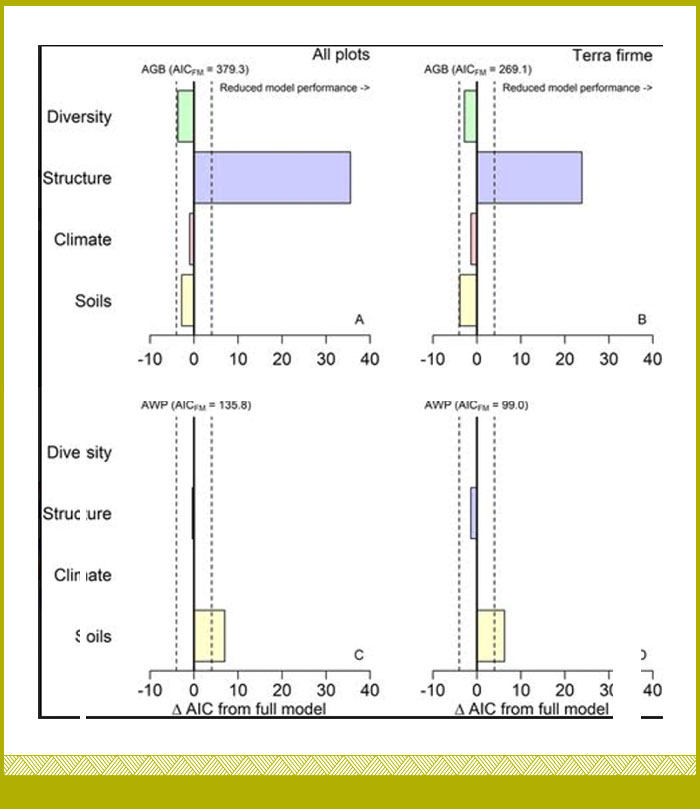
Encontramos también que los principales impulsores de la productividad de biomasa aérea (AWP) fueron las concentraciones de Fósforo (P) y el Magnesio (Mg) en el suelo, y la AWP no tuvo efectos significativos de los índices de diversidad. Estos hallazgos enfatizan la necesidad de incluir los principales cationes del suelo distintos del Nitrógeno (N) y el Fósforo (P) (por ejemplo, Mg) en los estudios experimentales para mejorar nuestra comprensión sobre el grado en que la fertilidad del suelo puede modular los aumentos en la productividad de biomasa debido al cambio climático. Los bosques de tierra firme presentaron mayores reservas de biomasa que los bosques inundados, pero ambos tenían una productividad similar. Nuestros resultados proporcionan evidencia limitada de los fuertes efectos de la diversidad vegetal en AGB o AWP. Nuestros resultados llaman hacia la cautela sobre las generalizaciones de las iniciativas basadas en la naturaleza que tienen como objetivo preservar la diversidad sobre la base de la maximización de las reservas de carbono y la productividad, debido a la naturaleza compleja de los procesos que controlan la acumulación de carbono y los flujos de carbono en éstos bosques (Castaño et al 2024 https://doi.org/10.1111/ecog.06693)
Ecography - 2024 - Castaño - Contrasting drivers of aboveground woody biomass and aboveground woody productivity in lowland
Householder et al. 2024
Nuestro colega John Ethan Householder (http://orcid.org/0000-0002-5959-3504) del Instituto de Tecnología Karlsruhe de Alemania, lideró un estudio de los bosques inundables de toda la Amazonia (https://doi.org/10.1038/s41559-024-02364-1); el sistema de bosques inundables más grande y biodiverso de la Tierra.
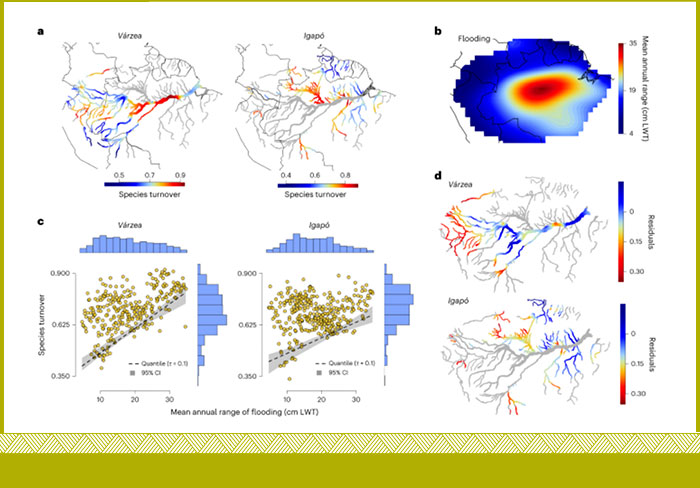
Nuestra comprensión de la composición de sus especies y de cómo ésta puede diferir de los tipos de bosques circundantes sigue siendo demasiado limitada, sobre todo a medida que los cambios en los regímenes de inundación comienzan a remodelar las comunidades arbóreas de las llanuras inundables y las funciones ecosistémicas críticas que sustentan. Aquí abordamos esta brecha mediante una mirada espacialmente explícita a los patrones de recambio de especies arbóreas y la especialización ecológica de los bosques de las llanuras inundables en toda la pan-Amazonía (Householder et al 2024). Mostramos que la mayoría de las especies arbóreas amazónicas pueden habitar en las llanuras inundables, y que alrededor de una sexta parte de la diversidad arbórea amazónica está ecológicamente especializada en las llanuras inundables.
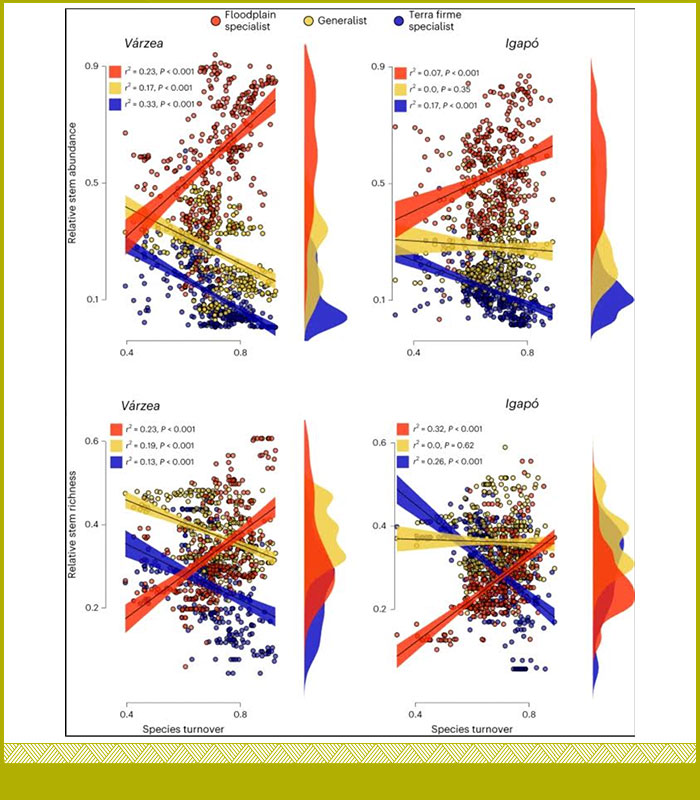
El Dr. Householder y colaboradores proporcionan una visión espacialmente explícita de la especialización ecológica de las comunidades de bosques inundables y exponen la necesidad de la integridad hidrológica de toda la cuenca para proteger la diversidad arbórea de la Amazonía y su función (Householder et al 2024 https://doi.org/10.1038/s41559-024-02364-1).
Nuestro colega Bruno Garcia Luize (https://orcid.org/0000-0002-8384-8386) de la Universidad Estatal de Campinas – UNICAMP, Brasil, lideró un estudio de la composición filogenética de los árboles en la pan-Amazonia (https://doi.org/10.1111/jbi.14816). Usando datos de abundancia de 5082 especies de árboles provenientes de 1989 parcelas, generamos una ordenación evolutiva para evaluar como varia a composición filogenética en toda la Amazonía. En los tipos de bosque de tierra firme y várzea, la composición filogenética varía según la región geográfica, pero los tipos de bosque igapó y de arena blanca conservan una rama evolutiva única independientemente de la región (Luize et al 2024 - https://doi.org/10.1111/jbi.14816). En general, encontramos que la química del suelo, el clima y la topografía explican el 24% de la variación en la composición filogenética, y el 79% de esa variación está estructurada espacialmente (en general para los efectos espaciales/ambientales combinados). La composición filogenética también muestra patrones espaciales sustanciales no relacionados con las variables ambientales cuantificadas. Un mayor número de linajes fueron indicadores significativos de las regiones geográficas más que del tipo de bosque (Luize et al 2024 - https://doi.org/10.1111/jbi.14816)
El Dr Cooper (http://orcid.org/0000-0002-2466-7710) de la University College London, UK lideró un estudio sobre los patrones de abundancia de las especies arbóreas más comunes a lo largo de los trópicos del mundo (https://doi.org/10.1038/s41586-023-06820-z). Utilizando datos de inventario de 1.003.805 árboles 1.568 parcelas ubicadas en bosques tropicales estructuralmente intactos en África, Amazonia y el sudeste asiático.
Estimamos que el 2,2%, 2,2% y 2,3% de las especies comprenden el 50% de los árboles tropicales de estas regiones, respectivamente. Extrapolando a todos los bosques tropicales de dosel cerrado, estimamos que solo 1.053 especies comprenden la mitad de los 800.000 millones de árboles tropicales de la Tierra. A pesar de las diferentes historias biogeográficas, climáticas y antropogénicas, encontramos patrones notablemente consistentes de especies comunes y distribuciones de abundancia de especies en todos los continentes. Esto sugiere que los mecanismos fundamentales de ensamblaje de comunidades arbóreas pueden aplicarse a todos los bosques tropicales (Cooper et al 2024 - https://doi.org/10.1038/s41586-023-06820-z). Los análisis muestran que es probable que las especies más comunes pertenezcan a una lista manejable de especies conocidas, lo que permite realizar esfuerzos específicos para comprender su ecología.
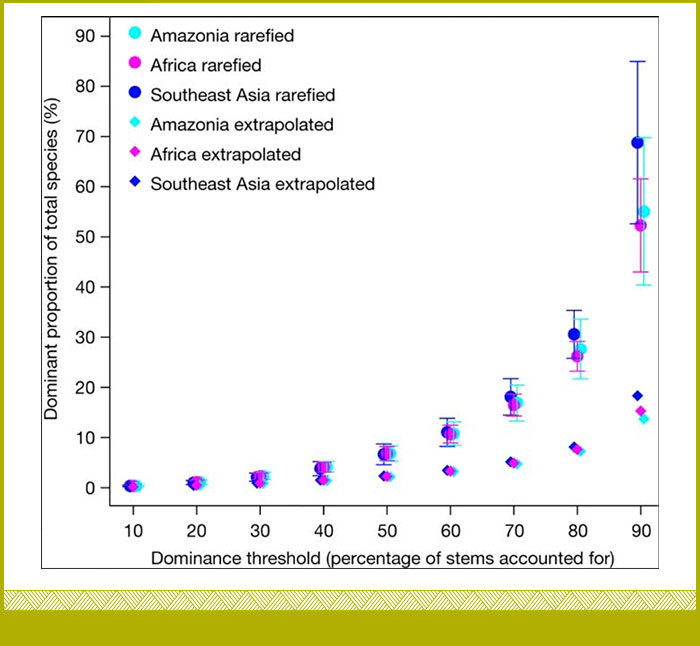
Aunque no restan importancia a las especies raras, nuestros resultados abren nuevas oportunidades para comprender los bosques más diversos del mundo, incluida la modelización de su respuesta al cambio ambiental, centrándose en las especies comunes que constituyen la mayoría de sus árboles (Cooper et al 2024 - https://doi.org/10.1038/s41586-023-06820-z).
2023
Bajo el liderazgo de nuestro colega Hans ter Steege (http://orcid.org/0000-0002-8738-2659) del Naturalis Biodiversity Center de Holanda, se usó información de 2.046 parcelas de árboles de bosques de la Amazonía y mapeamos la diversidad de especies arbóreas y la riqueza de especies arbóreas con una resolución de 0,1 grados, e investigamos los motores de la diversidad y la riqueza (https://doi.org/10.1038/s42003-023-05514-6). Utilizando la ubicación, estratificada por tipo de bosque, como predictor, nuestro modelo espacial, proporciona el mapa más preciso de la diversidad arbórea en la Amazonía hasta la fecha, explicando aproximadamente el 70% de la diversidad arbórea y la riqueza de especies (ter Steege et al 2023 https://doi.org/10.1038/s42003-023-05514-6).
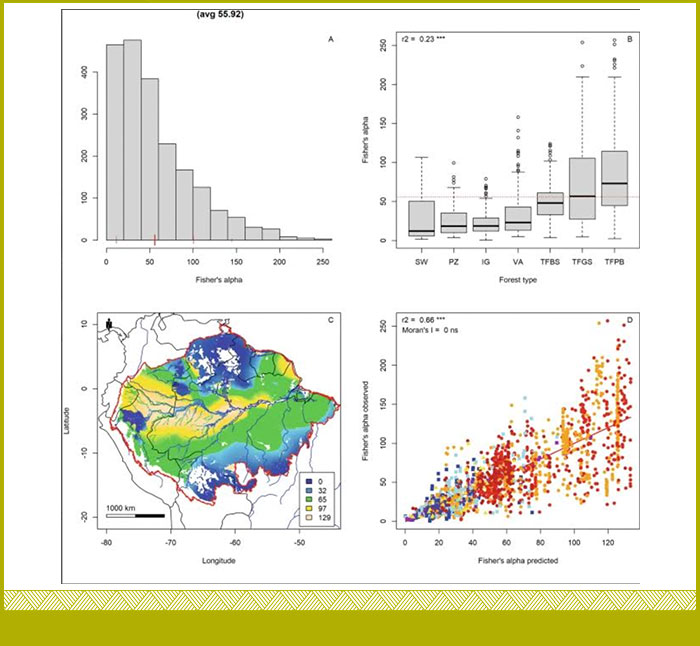
Histograma alpha-diversidad de árboles (ter Steege et al 2023)
Las grandes combinaciones suelo-bosque determinan un porcentaje significativo de la variación en la riqueza de especies arbóreas y la alfa diversidad arbórea en la Amazonía. Sugerimos que el tamaño y la fragmentación de estos sistemas impulsan sus patrones de diversidad a gran escala y, por lo tanto, la diversidad local. Un modelo que no utiliza la ubicación, el déficit hídrico acumulado, la densidad de árboles y la estacionalidad de la temperatura explica el 47% de la riqueza de especies arbóreas en el bosque de tierra firme en la Amazonía. El área del Escudo de Guyana tiene una riqueza de especies arbóreas menor a la esperada por nuestros modelos. Proporcionamos metadatos extensos de la parcela, incluidos los resultados de densidad de árboles, diversidad alfa de árboles y riqueza de especies de árboles, y mapas cuadriculados con una resolución de 0,1 grados (ter Steege et al 2023 https://doi.org/10.1038/s42003-023-05514-6).
2022
Correa et al 2022
Liderado por nuestro colega de la Universidad de los Andes Diego F Correa (https://orcid.org/0000-0002-6510-4906), colaboramos con entidades nacionales e internacionales para comprender la distribución geográfica de los síndromes de dispersión de las semillas (endozoocoria, sinzoocoria, anemócora e hidrocoria[1]) más comunes en los árboles de la pan-Amazonía (Correa et al 2022- https://doi.org/10.1111/geb.13596). Examinamos si la abundancia proporcional de estos síndromes de dispersión podría explicarse por la disponibilidad de agentes de dispersión y / o por la disponibilidad de recursos para construir frutos dispersados por animales (zoocoria) (hipótesis de disponibilidad de recursos) (Correa et al 2022 - https://doi.org/10.1111/geb.13596).
Encontramos que el síndrome de dispersión por aire (anemocoria) se asoció significativamente y positivamente con la velocidad media anual del viento. La naturaleza está finamente acondiciona, para que, en época de mayores vientos estén dispuestos los frutos que así lo requieran para dispersarse. Mientras que la dispersión por agua (hidrocoria) fue significativamente mayor en los bosques inundables, lo que demuestra la adaptación de las plantas a estos ambientes. En los bosques de tierra firme se encontró una menor disimilitud en los síndromes de dispersión, como resultado de un mayor predominio de las especie adaptadas a que sus frutos sean consumidos y dispersados por animales (endozoocoria). La disponibilidad de recursos para la construcción de frutos zoocorosos parece una explicación poco probable para la distribución de los modos de dispersión en la Amazonía. La asociación entre los frugívoros y la abundancia proporcional de los dispersados por animales requiere más investigación, ya que el reclutamiento de árboles no solo depende de los vectores de dispersión (Correa et al. 2022 - https://doi.org/10.1111/geb.13596).
[1] Endozoocoria: endo = adentro, zoo =animal, semillas comidas por un animal y dispersadas al pasar por su tracto digesntivo, sinzoocoria: frutos grandes que el ave/roedor debe transportar por fuera de su tracto, anemócora: fruto dispersado por el viento, normalmente con alas e hidrocoria: dispersado por el agua)

Marca-Zevallos et al 2022
Liderado por nuestro colega del INPA y la Univ. Nacional de San Antonio Abad del Cusco, Manuel Marca-Zevallos (https://orcid.org/0000-0001-9428-7441), en donde usamos 443 parcelas de toda la Amazonía, y registramos 210,801 individuos de 3,527 especies (https://doi.org/10.1111/ecog.06125).
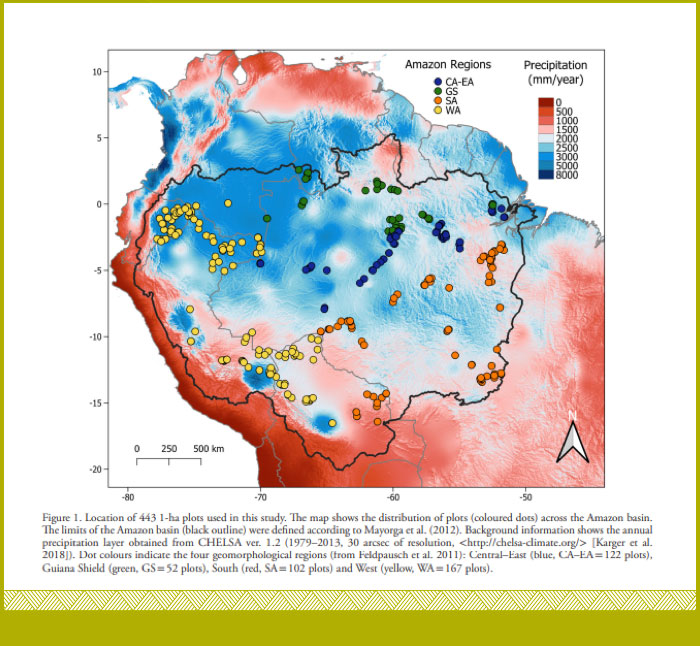
Con Manuel y colaboradores, encontramos un gran efecto de las condiciones hidrológicas locales sobre la diversidad de árboles (https://doi.org/10.1111/ecog.06125), siendo más evidente en los bosques más húmedos, donde la diversidad aumenta hacia lugares con suelos bien drenados. Encontramos que la media ponderada del valor de la densidad de la madera por comunidad aumentó hacia suelos mejor drenados en el sur y oeste de la Amazonía. Nuestros resultados sugieren que los gradientes hidrológicos locales filtran especies, influyen en la diversidad y composición de los bosques amazónicos (Marca-Zevallos et al 2022 - https://doi.org/10.1111/ecog.06125). Lo cual evidencia la gran complejidad de los éstos bosques. La composición de las especies de árboles cambió a lo largo de los gradientes hidrológicos locales del suelo en la Amazonía centro-oriental, occidental y meridional, y esos cambios se correlacionaron con cambios en la densidad media de madera de las parcelas. Nuestros resultados sugieren que los gradientes hidrológicos locales filtran especies, influyendo en la diversidad y composición de los bosques amazónicos. En general, este estudio muestra que el efecto de las condiciones hidrológicas locales es generalizado, se extiende sobre amplias regiones amazónicas, y refuerza la importancia de tener en cuenta la topografía y la hidrología locales para comprender mejor la probable respuesta y resiliencia de los bosques al aumento de la frecuencia de eventos climáticos extremos y el aumento de las temperaturas (Marca-Zevallos et al 2022 - https://doi.org/10.1111/ecog.06125).













